John W. Pelley és Edward F. Goljan könyvéről nem azt állítom, hogy a legjobb biokémia könyv, ami valaha is megjelent, amit viszont ki merek jelenti, hogy a legjobbak egyike és a legpraktikusabb a magyar piacon.
Nos, természetesen vannak témák, amit nem lehet és nem is szabad rövidíteni egy átfogó jellegű könyvben, viszont a biokémia alighanem ilyen. Természetesen nem pótolja a jó öreg, folyamatosan frissített Orvosi biokémia és a hasonlóképpen megkerülhetetlen Orvosi pathobiokémia kötetet, de nem is arra találták ki. Bármiről is legyen szó, nem az a lényeg, hogy mennyit tudunk valamiről megtanulni, hanem az, hogy mennyit tudunk megjegyezni, amire viszont ez a kisebb biokémia kötet alkalmasabbnak tűnik. A didaktikai összeszedettség mellett az apró klinikai kitekintések és a gyakorlatias tárgyalás miatt lopta be magát ez a könyv annak idején a szivembe.
Azt hiszem, hogy manapság alig van kutató, akinek tényleg fejből kellene tudnia például egy-egy biokémiai folyamatnál az összes izomerizációs lépést [a kulcs lépéseket annál inkább!], az természetesen elvárható, hogy egy-egy lépésre azonnal rá tudjunk keresni a neten, ha az előkerül.
A könyvet egy, az ismeretek megértését elmélyítő, gondolkodtató teszt zárja, amire a helyes válaszok - nagyon helyesen - pusztán lexikális tudással nem tippelhetőek meg, viszont természetesen van egy bőséges magyarázó rész a helyes megoldásokhoz.
Nemrég a Genomika Blogon olvastam, hogy az NCBI megszünteti a Sequence Read Archive genomi szekvenciaadatokat kiszolgáló adatbázisát. Az NCBI nem indokol, inkább ajánl helyette 5 másik adatbázist.
A Genomika Blogon viszont azt olvashatjuk, hogy az NCBI esetleg nem tudott elég háttértárat beboltolni vagy a sávszélesség biztosítása fájt túl sokba a bioinformatikai óriásszervezetek. Természetesen ez is lehetséges, én viszont – a Sequence Read Archive pontos felépítését nem ismerem ugyan – kevésbé prózai tényezőket tartok valószínűnek, ami miatt az adatbázist a régi formájában már nem tartják fenn. Teljes bizonyossággal pedig, senki sem tudná megmondani, hogy miért is szüntették meg, erről fogok most írni.
Bármilyen informatikai projekt vagy működő rendszer később elcsúszhat olyan dolgokon is, amit alapos előre tervezéssel meg lehetett volna előzni, utólag viszont senki sem vallaná, hogy hibázott. Az egyik, talán legfontosabb, hogy a rendszer működtetéséhez szükséges hardver- és szoftverkomponensek legyenek egymástól annyira függetlenek, amennyire csak lehet (régebben beállított rendszereknél ez annyira nem természetes), ezen kívül mindig arra a megoldásra kell hajlani, ami olcsóbb. Hogy a webprogramozás köréből említsek egy nagyon egyszerű példát – akármerre is navigáltok a neten, sokszor a böngésző címsorában látható, hogy a cím php-ra végződik, ami most annyit jelent, hogy az adott dinamikus webhelyet szerveroldalon egy PHP technológiában megvalósított szoftver generálja ki, majd küldi vissza a böngészőprogramnak a ténylegesen megjelenítendő, ún. kliensoldali kódot. Nos, az ún. PHP-interpreter, ami a PHP nyelven megírt programot futtatja a szerveren, többféle is lehet, ezek közül a két legelterjedtebb a PHP Community által kiadott, teljesen ingyenes PHP interpreter, a másik pedig a Zend Technologies által kínált Zend Web Application Server, amiért viszont kemény pénzeket kell fizetni* függően többek közt attól, hogy az IT staff melyik terméktámogatási szint mellett dönt. Abban az esetben, ha egy rendszert felhúznak egy Zend motorra, majd a későbbiekben kiderül, hogy a PHP Community ingyenes PHP motorja több szempontból kedvezőbb, legrosszabb esetben egy éjszaka alatt a szerver(ek)en ki lehet cserélni az egyik motort a másikra anélkül, hogy ez komolyabb kiesést eredményezne. De gondoljunk bele abba, hogy ha egy internetszolgáltató webhosting szolgáltatásáról van szó, és a korábbi Zend motor több tízezer dinamikus webhelyet szolgál ki egy időben! Vagy éppenséggel a világ legnagyobb közösségi hálója, a Facebook alatt próbálnák kicserélni a Zend motort, miközben több millió felhasználó használja egyidejűleg! (A FB természetesen a kezdetektől a PHP Community motorját használja.) Ilyen esetben, ha a szolgáltatás rövid kimaradása nem lenne elég ciki, azzal is számolni kell, hogy az új interpreter a PHP-kódokat esetleg nem ugyanúgy futtatja, mint a másik, így a teljes webszolgáltatás nem működik normálisan, ergo legrosszabb esetben a konfiguráció finomhangolása néhány órás rémálommá válhat, mire a rend helyreáll.
És ez csak egy egyszerű példa volt! Lehet szó olyan szoftverkomponensről is, aminek a használati jogát az NCBI olyan szerencsétlenül vásárolta meg, hogy arról nem tud átváltani egy újabban megjelent olcsóbb vagy teljesen ingyenes verzióra, horribile dictu egy ilyen fizetős komponensre épül más, a rendszer működéséhez elengedhetetlen szoftver. Azaz az egészet vérrel-verejtékkel a felismerhetetlenségig át kellene operálni, egy új szoftveres architektúrára váltás esetén.
Az adattárolás módjával kapcsolatos változtatás kevésbé játszhat be, mert egyik típusú adatbázisból egy másik típusú adatbázisba való átkonvertálás nem probléma, általában. De gondoljunk bele, hogy akár az Oracle és a MySQL, akár a MySQL és annak követhetetlenül sok klónja közt is mennyi apró eltérés van, amit mind figyelembe véve azt mondhatjuk, hogy egy extrém nagy rendszer esetén a programozót ki lehet vele kergetni a világból.
Ha pedig specifikus területet nézünk, azt látom, hogy a bioinformatikai szoftverek piacán a kompatibilitás megőrzése vagy fenntartása szinte az utolsó szempont, amiből szintén adódhatnak galibák egy átálláskor.
Azaz általánosságban elmondhatjuk, hogy bármilyen rendszer áthelyezhető ugyan akár teljesen eltérő technológiai alapokra is kisebb-nagyobb energiabefektetéssel, gyakorlatilag ilyen csak akkor történik, ha már minden kötél szakad. Én konkrétan tudok olyanról, hogy egy több millió ügyféllel rendelkező telekom szolgáltató csillagászati összegért beboltolt egy olyan szoftverrendszert, ami nap, mint nap a legváltozatosabb hibajelenségeket produkálja, a szoftverrendszer support csoportja mégis sokszor csak tüneti kezelést tud alkalmazni. A dolog pikantériája, hogy nyílt forráskódú rendszer ide vagy oda, abban ez esetben, ha a szóban forgó szolgáltató, mint ügyfél, megbízna egy szakit a rendszer feljavításával, amint valaki belekotorna a forráskódba, a méregdrága support-licensz elveszne.
Az általam kevéssé ismert szekvencia adatbázis esetén tehát jóval valószínűbb, hogy időben fellépő kompatibilitási problémák tömege jelent meg, amiknek a kiküszöbölése olyan égtelenül drága lett volna, hogy az NCBI célszerűbbnek tartotta más adatbázisaira fordítani azt a pénzt, amit átállításra fordítottak volna, ha a szolgáltatás továbbra is marad.
*több helyen kicsit szájbarágós lett a poszt, mielőtt valaki megjegyezné: igen, tudom, hogy a Zend-nek is vannak ingyenes megoldásai viszont ahhoz semmilyen terméktámogatás nem jár és – lévén, hogy nem nyílt forrású szoftverről van szó – azon túl, hogy egy-egy, az interpreter sajátosságából adódó beállítás kikutatása is jókora szopó lehet, biztonság szempontjából is rizikós, különösen nagy bioinformatikai rendszereknél, ami többek szerint a kiberterrorizmus 2-3. legnagyobb célpontjaivá váltak.
A személyre szabott genetika/genomika témában természetesen Meskó Berci blogját ajánlom, viszont ezúttal nekem is sikerült előpecázni valamit az internet susnyásából, ami érdekes lehet mindenkinek, aki egy kicsit kíváncsi rá, hogy milyen betegségekre számíthat későbbi élete folyamán nagyobb valószínűséggel.
Amit már most leszögeznék, hogy a hiedelemmel és a hatásvadász bulvárbárók süketeléseivel ellentétben, az ember sorsa genetikailag nem meghatározott, azaz a DNS-ben tárolt információ, ami meghatározza, hogy egyes génekből milyen típussal rendelkezünk, nem irányítója, hanem csak kiszolgálója a sejtnek, ezen keresztül a teljes szervezetnek is. A genotipusnak nagyon sok esetben egyszerűen nem következménye a fenotipus, ami tehát a genetikai tulajdonságaink kifejeződésének és a minket érő környezeti hatásoknak az interakciójából adódik össze.
Amikor genetikáról beszélünk, nagyon sok esetben csak valószínűségeket lehet mondani egy tulajdonsággal kapcsolatban, legyen az egy betegségre való hajlam, valamilyen anatómiai vagy élettani paraméter vagy olyan magas szinten szerveződő tulajdonságok, mint amilyenek az egyes személyiségjegyek.
Ma már a neten számos helyen lehet rendelni olyan teszteket, amik adott genetikai tulajdonságunkat vizsgálják. A móka viszonylag egyszerű: az adott cégnél csak be kell regisztrálni több-kevesebb összegért, mire ők küldenek egy kis flaskát, amibe csak bele kell köpni, aztán a készítmény már mehet is vissza postán a tesztet elvégző cégnek elemzésre.
Persze a teszteket végző cégek rendszerint megpróbálják túllicitálni egymást árban, na meg feature-ökben, most például a 23andMe mostani dobására lettem figyelmes: nem kevesebb, mint 95 különböző megbetegedésre valószínűsítő faktor kimutatását ajánlják 199 USD-ért, a teljes vizsgálható repertoár pedig a honlapjukon is fent van. A 23andMe még tovább ment azzal, hogy plusz néhány dolláros havidíjért a korábbi ügyfeleket tudják értesíteni az olyan kockázati tényezőkről, amikről korábban még nem tudták, hogy kockázati tényezők, viszont a korábban beküldött és már "felvett" egyéni DNS minta alapján felismerik azt, így tudnak értesítés küldeni róla, hiszen a jövőben felfedezésre váró hajlamosító gének helye vagy típusa akkorra már ismert lesz, azaz egybevethető korábbi, adatbázisban tárolt mintákkal.
Így elnézve a listát, aki valamelyest vágja, hogy miről van szó, inkább nyel egyet és azt gondolja magában, hogy jobb nem tudni, hogy milyen kórságok várnak rá nagyobb sansszal. Nos, szerintem mégiscsak jobb tudni, hogy hajlamos-e valaki a Hodking-limfómára, skizofréniára vagy éppen a heroin függőségre – viszont fontos, hogy csak akkor, ha ismeri mindezek környezetből jövő hajlamosító tényezőit és tud is ellene tenni.
Érkezik, amit legalább hónapok óta szinte biztosra lehetett tudni – a Facebook e-mail szolgáltatással áll elő. Na, hogy precíz legyek: fél-SMTP-szolgáltatással. Azért fontos kiemelni ezt, mert ahogy azon már korábban kihőbörögtem magam, jegyezzük meg: a RajVIP-en, prosztó-facebookon (WiW-en), MySpace-en és Facebook-on küldött levelek elektronikus levelek ugyan, de NEM e-mailek, a különbség pedig óriási. Ahogy arról is írtam már hébe-hóba, hogy komoly globális kockázatot jelent az, ha az elektronikus levelezés áthelyeződik a jó öreg, visszakövethető, elemezhető, digitálisan aláírható, hitelesíthető, titkosítható, azaz gyakorlatilag bármilyen RFC –ajánlást megvalósítani képes e-mail szolgáltatásról közösségi szolgáltatások üzenetküldő rendszereire.
A MySpace-en egyébként már régen létező, kicsit hasonló funkcióról van szó, így például a – sosem használt – molbiol@myspace.com címre érkező levelek is megérkeznek, ugyan csak a MySpace-en belül.
Tény, a netet, ha nem is a gagyi, de mondjuk úgy, a puritán módon egyszerű e-mail szolgáltatások uralják. Ha elkapsz valakit egy kétperces beszélgetésre, majd szóba kerül, hogy mi jut eszébe az e-mailről, ami eszébe fog jutni, a Gmail, Hotmail, Live, AOL, Yahoo, ha esetleg az illető az utóbbi egy évtizedet egy barlangban töltötte elzártan a civilizációtól, szóba kerülhetnek még olyan nemzeti szégyenfoltjaink, mint a Freemail vagy a WiW-es üzenetküldés. A felsorolt levelezőrendszerek egy részének ingyenes változatában egyébként a mai napig elérhetetlennek az olyan alapszolgáltatások, mint amilyen a POP3/secure POP3-elérés, az IMAP és még sorolhatnám, hiszen a felhasználók döntő része úgyis webes felületen keresztül levelezik, mintha más elérési forma a világon nem is létezne. És persze, amit tök természetesnek vesz az egység sugarú felhasználó, hogy az e-mail ingyenes, mert hát miért is fizetne érte. Mondjuk amiért nem fizet, kártyával, ott fizet az adataival, de ebbe most nem mennék bele. Pont tegnap csevegtem erről egy témában kevésbé járatos barátosnémmal arról, felhasználói igényektől és a felhasználó internetes műveltségétől függően még az e-mailezésnek is külön szintjei vannak, amik nagyjából a következő módon oszthatóak fel.
1.Valamilyen ingyenes mailbox, olyan alapon, mert „a másik is ezt használja”, annak minden előnyével és hátrányával. Érdekes megfigyelés, hogy amikor a felhasználók felhasználói nevet választanak, tipikusan nem aszerint döntenek, hogy azt hogyan tudják majd levelezőpartnereik minél kisebb eséllyel félrecímezni őket, fejben tartani, azaz a cím minél elegánsabb legyen mindenféle debil karakterek és számjegyek nélkül. Sokkal inkább az alapján döntenek, hogy önmaguk számára könnyen megjegyezhető címet válasszanak, aminek eredményeként néha ritka szánalmas címeket tudnak kisütni. Mindenesetre a legalja bizonyára az a kategória, amikor valaki egy névrészletét fogalmazza a felhasználói névbe és ezt még aláhúzás jellel, és születési évvel is súlyosbítja. Na ettől még egy fantázianév is kevésbé lenne ciki.
2.Saját internetszolgáltató által adott mailbox, ami ugyan „ingyenes”, viszont a mezei netszolgáltatónak vagy mobilszolgáltatónak nyilván nem érdeke minél professzionálisabb e-mail rendszert kialakítania. Másrészt ha nagy szolgáltatóról van szó, ott is bejöhetnek a képbe a felhasználói név formátumával kapcsolatos megszorítások, úgy, mint a júzernév minimális hossza, valamint az az „apróság”, hogy a saját keresztnév már úgysem adható meg, mert arra valaki korábban már úgyis lecsapott. Hacsak nem olyan nagyon hülye névvel áldotta/verte meg a sors a felhasználót, mint mondjuk Dzsindzser, Szüvellő, Szederina, Andzseló vagy Fernanó. /*A fenti neveket vastagbőrék blogjáról puskáztam */
3.Ha valaki saját domain-nevet, DNS-szolgáltatást használ önmagában vagy tárhellyel, esetleg virtualszerverrel, saját DNS-konfig esetén olyan levelezőrendszert választ, amilyet csak akar, kis trükközéssel akár gyakorlatilag korlátlan e-mail címet csaphat saját domain-neve alá. Így például a Molbiol Inside elsődlegesen a Google Applications-t [GHS, Google Hosted Services] használja, ami ár-érték arányban verhetetlen, hiszen a Standard csomagjuk totál ingyenes. Ha pedig az abban foglalt domainenkénti 50 regisztrálható felhasználói név nem lenne elég, egy trükkel megoldható a további bővítés anyagi ráfordítás nélkül. Ami még fontos lehet, hogy a GoogleMail motor által hajtott pici vagy annak tűnő magán-levelezőrendszerek egy csomó megszorítástól eltekintenek a Gmail és a Google szolgáltatásainak megszorításaihoz képest.
4.Aki nem sajnál rá évente néhány garast, érdemes lehet egy kis tájékozódást követően előfizetni valamilyen prémium e-mail szolgáltatásra. Az előfizetés előtti tájékozódás mindenképp fontos, hiszen attól, hogy valami fizetős, még nem feltétlenül annyira minőségi is, pontosabban nem biztos, hogy mindent tudni fog, ami kellhet. Személy szerint összeesküvésszagú ostobaságnak tartom azokat a feltételezéseket, hogy az extra-biztonságos prémium e-mail szolgáltatások, mint a hushmail.com vagy a safe-mail.net ilyen-olyan nemzetbiztonsági szolgálatok megfigyelő rendszerei lennének, amiknél kimondottan azoknak az ügyfeleknek a forgalmát stírolnák ráérős fekete öltönyös bácsik, akik valami miatt ilyen e-mail szolgáltatásra fizetnek elő.
-A hushmail.com filozófiájához hozzá tartozik, hogy a levelek még csak ideiglenesen se nagyon jelenhessenek meg vagy tárolódjanak titkosítatlan formában, így a POP3 letöltési lehetőség szándékosan nincs is benne, ami a levelek elérését például mobileszközről számomra elfogadhatatlanul kényelmetlenné teszi. /*secure IMAP van ugyan, de annál még a mobilwebes elérés is rugalmasabb és kevésbé idegesítő*/. Ezen kívül a hushmail-en minden apróságért pluszban fizetni kell, nem is keveset, a néhány hónappal ezelőtti több, mint fél napos leállás pedig röhejessé teszi azt, ahogy a Hushmail bombabiztos szolgáltatóként sztárolja magát.
-safe-mail.net – szivem csücske, de tényleg. Az ingyenes változatban már alapértelmezés szerint majdnem minden feature elérhető, ami a fizetősben, ugyan az egész csupán 3 MB-os tárhellyel rendelkezik, ami számomra mellékes, hiszen secure-POP3-on keresztül cibálom át az oda érkező leveleimet automatikusan saját levelezőrendszerre, így az a 3 MB nekem még sosem telt be. A több száz professzionális – értsd úgy: átlag felhasználónak sosem kellő – extra közül, úgy, mint a digitális aláírás, igény szerinti titkosítás, az utolsó 500 belépés és belépési kísérlet IP-jének és idejének lekérdezhetősége mellett személyes kedvenc a SafeBox-funció. Ezzel titkosított levél küldhető olyan levelezőrendszert használó felhasználónak is, aki egyébként a titkosított leveleket egyáltalán nem tudná kezelni. A címzett egy linket kap és egy jelszót, aminek megadása után tudja olvasni, amit küldtem neki, ráadásul a levél életideje beállítható, azaz mennyi ideig maradjon a levél olvasható a megérkezéstől és az elolvasástól számítva.
-A fullextrás runbox.com az utóbbi időben egy másik nagy szerelmemmé. Gyors is, okos is, az agyonmajmolt Gmail taknyosorrú hülyegyerek hozzá képest és ami fontos – olcsó! Ingyenes változata természetesen a Runboxnak is van, abban nem minden funkció jelenik meg, a legkisebb fizetős csomaghoz képest.
A legelegánsabb persze még mindig a céges vagy egyetemi cím, ami a hiedelemmel ellentétben nem szűnik meg akkor, ha a felhasználó munkahelyet vált, már ha a rendszergazdával jóban van. A bökkenő csak az vele, hogy egyrészt még a legprofibb egyetemi levelezőrendszerek sem képesek adni a prémium levelezőszolgáltatások által nyújtott extrákat, hiszen nem is arra tervezték őket, másrészt néhányan nem szivesen küldenek személyes tartalmú levelet hivatalos címre. Nekem mondjuk mind a 4-5 ilyen-olyan egyetemhez tartozó címemre érkező levelek azonnal pattannak is át máshova, még mindig a legelegánsabb megoldás, ha már az e-mail címről látszik, hogy a tulajdonosa tartozik valahová, minél pofásabb a domain, annál jobb.
Összefoglalva az eddigieket, azt tudnám írni, hogy az elektronikus levelezésnek ilyen aspektusból még nem terjedt el széles körben a kultúrája, ami viszont megállapítható egy bedemártonos szóhasználattal, hogy az egyetemi, céges cím menő, a prémium mailszolgáltatás menő annak, aki tudja, hogy az micsoda, a kommersz mailcím használata nem menő, a közösségi szolgáltatások belső üzenetküldője az elkerülhetetlen szükséges rossz, nagyon kommersz ingyenes magyar mailszolgáltatók használata sokszor vérciki módon nem menő. Ami a Facebook e-mailt illeti, borítékolhatóan béget majd a nyáj a privacy miatt, de bejósolhatóan majdnem mindenki használja majd.
Amit nem vesz észre szinte senki – a Facebook az erőforrások-megtakarításán keresztül egy vagonnyi pénzt spórol majd meg azzal, hogy az e-mail notification-öket nem külső mailszolgáltatóhoz kell küldenie, hanem elég lesz házon belül kézbesíteni.
Szép dolog is a biokémia. A probléma csak az vele, hogy ha valaki nem kimondottan valamelyik biokémiai folyamattal foglalkozik, garantáltan el fogja felejteni azt, amit annak idején a középiskolában vagy az egyetemen tudnia kellett. A lényeg persze nem is ebben áll, hanem abban, hogy a biokémiai folyamatok lényegét jegyezze meg a hallgató. Ezzel viszont nem is lesz probléma, hiszen ha az újonnan megszerzett ismeretek lényege kapcsolódik addigi természettudományos ismeretei logikai rendjébe, akkor az emlékezetében megtapad, ha nem, akkor nem. Értelemszerűen az általános biokémiai tárgyú könyveknél tényleg a méret a lényeg: minél kisebb, annál használhatóbb.
Az érdeklődő emberben biztosan felmerült már az a kérdés, hogy mennyire is bonyolult egy élő sejt működése? Röviden megválaszolva: nagyon. Azok az anyagcsereutak, amik leírják, hogy milyen anyag milyen más anyaggá alakul át egy-egy enzim hatására, nem meglepő módon hálózatot alkotnak. Jópár enzim (és annak termékei) közül némelyiknek a defektusa vagy hiánya teljesen összeegyeztethetetlen az élettel, megint más esetben sokáig azt hitték, hogy összeegyeztethetetlen az élettel, aztán kiderül, hogy mégsem. Itt konkrétan Falusék hisztamin knock-out egereire gondolok, azaz olyan génmódosított egérkékre, amiknek a sejtjei egyáltalán nem termelnek hisztamint, amit többek közt az immunrendszeri folyamatok egyik főszereplőjének tartottak, az ilyen hisztaminhiányos egerek mégis köszönik szépen, jól vannak. Más-más enzimhibák okolhatók súlyosabb és kevésbé súlyos anyagcsere betegségekért, aminél tehát „csak” arról van szó, hogy az egyik vegyület nem tud átalakulni egy másik vegyületté.
Anyagcsere-folyamatokból nagyon sok van, ezek hálózatának magával ragadó bonyolultságát a Svéd Bioinformatikai Intézet egyfajta fanyar humorral tárja az érdeklődők elé. A Roche még évekkel ezelőtt készített egy posztert, amin feltüntette gyakorlatilag az összes biokémiai reakciót, amit a biokémia könyvek emlegetnek (a makromolekulákkal kapcsolatos reakciókat kihagyták). A teljes ábra természetesen hatalmas nagyra sikerült irritálóan apró betűkkel. Ezt a posztert a svédek, mint egy GPS-térképet, interaktívvá tették, az így nyert ábra pedig normális ember számára első blikkre leginkább egy mikroprocesszor kapcsolási rajzának tűnik. Igen ám, viszont elég egyetlen kattintás az ábra valamelyik részére, máris látható válik a kiválasztott területen felrajzolt konkrét reakció bizarrabbnál bizarrabb vegyületnevekkel, természetesen a reakcióban részt vevő enzim megjelölésével turbósítva. Nem újdonság, de azért jópofa.
Már napokkal ezelőtt elhatároztam, hogy dobok egy posztot korunk egyik népbetegségéről, a daganatos megbetegedésekről. Onkológus ugyan nem vagyok, az már előre látszott, hogy nem leszek könnyű helyzetben, ha erről olyan posztot szeretnék írni, amiben a tumoros megbetegedések alapvonásait szakszerűen, mégis könnyen elfogadható nyelvezettel próbálom tálalni.
A rák, mint olyan, gyűjtőfogalom, több száz különböző típusú daganattípus van kialakulása, helye, áttétképzésre való hajlama és egyéb tulajdonságai alapján. Éppen ezért társaságban a félműveltség tuti biztos jele „a rák ellenszeréről”, „a rák gyógyításáról” úgy beszélni, mintha egy jól körülhatárolt kórságról lenne szó, holott sokszor mégis ez tapasztalható.
Tehát több száz, talán több ezer eltérő megbetegedésről van szó ugyan, ezeknek vannak markáns, közös alapvonásai, ami alól természetesen mindig akad kivétel.
Ami általában eszébe jut az újságolvasó embernek a tumoros megbetegedésekről, az az, hogy olyan sejtburjánzásról van szó, aminek eredete egyetlen sejt megkergüléséhez vezethető vissza. Többek véleménye szerint testünkben naponta több száz olyan tumorsejt keletkezik, amit ha nem fékezne meg azonnal az immunrendszer, az valamilyen gyors lefolyású daganatos megbetegedést alakítana ki. Még ha egy-egy hírportál be is számol valamilyen új hatóanyag felfedezésről, aminek a bevetésével hatékonyabban lehet segíteni a betegeken, mint korábban, akkor sem esik szó arról, hogy mi is váltja ki molekuláris biológiai szinten azt a folyamatot, aminek során egy daganat életveszélyessé vagy halálossá válik.
Ideális esetben a sejtek csak annyiszor és ott osztódnak és differenciálódnak, ahol osztódniuk kell, ezt saját genetikai programjukon kívül meghatározzák a környezetükből érkező jelek is, amit ún. szignál-transzdukciónak nevezünk. A sejt kaphat a környezetétől olyan jelet, ami arra utasítja, hogy osztódjon, de bizonyos esetekben egyenesen egy arra vonatkozó precíz tervet kell véghez vinnie, hogy kinyiffantsa önmagát (nekrózis, apoptózis). A néhány sorban leírt folyamat millió meg egy helyen meghibásodhat, ekkor a sejt kóros proliferációja – osztódása – veszélyezteti azt a szövetet, így szervet, amiben van, ezen kívül áttétet képezhet. A metastatis – áttétképzés – folyamán egy teljesen más szövetben megjelenő tumorból sejt(ek) jut át egy másik szövetbe, majd ott is kontrollálatlanul osztódni kezd, esetleg hibridizál eredetileg egészséges sejtekkel is, így a tumor a első megjelenésén kívül más szövetet is megtámad.
Szignál-transzdukció, avagy a lényeg – így kommunikál a sejt saját környezetével, majd enzimek így továbbítják az információt és utasítják a sejtet, hogy mit kell tennie éppen
Az összes rákos sejtre egyaránt igaz, hogy kromoszomális szinten vagy DNS-szinten rendkívül instabil. Ha a természetben genetikai szempontból valami instabil, hajlamos a mutációra, az egyben azt is jelenti, hogy olyan utódjai is meg fognak jelenni, ami a többiekhez képest előnyösebben alkalmazkodik a környezetükhöz. Nincs ez máshogy a rákos sejtekkel sem, amik amellett, hogy genetikailag nagyon variábilisak – egy bizonyos tumortípuson belül is – kikerülik a mitotikus ellenőrzőpontokat, azaz a sejtciklusnak azon pontjait, amiben különböző ellenőrző mechanizmusok azt vizsgálják, hogy az osztódás után újonnan létrejött sejt normálisan működik-e, azaz végezheti dolgát vagy éppenséggel halál rá, mert veszélyes. Tehát ez az, ami a rákos sejteknél hiányzik.
Az mára adatbázisokon keresztül néhány kattintással elérhető, hogy milyen defekt milyen tumor megjelenését valószínűsíti vagy teszi biztossá. Az emberi genom ugyanis számos ún. proto-onkogént tartalmaz, ami olyan génváltozat, ami önmagában nem bolondítja meg a sejtet, viszont valamilyen hatásra aktívvá válva igen. Az aktívvá válás oka - tumortípusonként - lehet az, hogy áthelyeződik a kromoszómán belül, felsokszorozódik, azaz több példány is megjelenik belőle egymás után, esetleg egyetlen nukleotid megváltozik a bázissorrendben (pont mutáció) történik vagy egy retrovírus fertőzés következményeként lesz a gén kifejeződése rendellenes.
A proto-onkogének egyébként eredeti feladatuk szerint szinte akármilyen gének lehetnek. A T-sejtes lymphoid leukemiában a TAL-1 gén helyeződik át egyik kromoszómáról a másikra, míg a Burkitt-limphomában az eredetileg is jelen lévő myc gén kerül a szintén eredetileg is jelen lévő, egyik immunoglobulin fehérje felépítéséért felelős gén elé.
Ismét más esetben egy eredetileg is jelen lévő génben olyan pontmutáció jelenik meg, ami miatt a sejtmembrán belső oldalán folyamatosan mitogén jelátviteli szignálfehérje jelenik meg, azaz a sejt osztódását nem a környezete szabályozza, hanem mintegy önmagát utasítja folytonos osztódásra.
Amiről még nem ejtettem szót, a tumor-szupresszor gének, amik a sejtek differenciálódásának és osztódásának gátlásában kulcs fontosságúak. Egy-egy ilyen gén defektje vagy teljes kiesése játszik be sokszor egy-egy tumortípus létrejöttében.
Első merítésre, néhány tankönyvi példa:
A legismertebb ilyen tumorszupresszor a p53, ami a DNS adott, specifikus helyeihez kapcsolódik, így a transzkripciót lehetetlenné teszi, így meggátolja bizonyos gének átíródását. A p53 bizonyos mutációinál egyszerűen ez a mechanizmus kiesik, holott ennek a génnek a normális működése lenne az egyik legfontosabb ellenőrzőpont a sejtosztódás azon részében, amikor eldől, hogy a sejt nem jelent-e veszélyt a környezetére. A normális típusú p53 aktiválódik több eltérő típusú DNS-károsodás, de például az osztódási orsó sérülése esetén is, ami a sejtosztódás során újonnan megjelenő kromoszómák mozgatásáért felel.
Nem világos, hogy hogyan, de a sejtet szintén veszélyeztető ionizáló sugárzás esetén az ATM gén is a p53-mal lép kölcsönhatásba, így felügyelve az esetlegesen károsodott sejtet. Ahogy az olvasó kitalálhatta, ennek a génnek a működésképtelensége szabad utat enged a súlyosan sérült sejtek osztódásának is.
A Nijmegen kromoszóma törékenységi szindróma esetén az NBS nevű génnek jelenik meg olyan változata, ami az ionizáló sugárzás okozta DNS-töréseket képtelen felismerni, holott ez lenne a dolga, emiatt többek közt a betegek az ionizáló sugárzásra különösen érzékenyek lesznek.
Az NF1 és NF2 gének elváltozásakor 1-es és 2-es típusú neurofibromatosis alakul ki, aminek eredményeként a szignáltranszdukció lesz dekfektes, így kialakítva az egyik legelvadultabb tumortípust.
A neurofibromatosis egy extrém méreteket is ölthet, főleg ha a beteg pl. vallási okokból nem enged korábbi orvosi beavatkozást
A VHL-génnel kapcsolatban figyelték meg, hogy az angiogenezist, azaz az érképződést gátolja a szövetekben, márpedig a nagyobb tumorok saját érrendszerrel rendelkeznek, így ennek a génnek a defektje is hozzájárul ahhoz, hogy a tumor saját érrendszert alakítson ki önmagának. Nem mellékesen ugyanez a gén normális esetben általánosan a sejtciklusból való kilépésért felelős, ami egyszerűen nem történik meg, ha hibás.
Ahogy azt az elmúlt néhány évtizedben felismerték, az élő sejt mindent megtesz annak érdekében, hogy osztódáskor a DNS tökéletesen másolódjon, ha a másolásban mégis hiba következik be, a hibát egy ún. repair enzimrendszer kijavítsa a hiba típusától függően. A DNS-hélixében adenin bázissal szemben csak timin, míg guaninnal szemben csak citozin lehet, mégis előfordul, hogy az újonnan létrejövő DNS-láncban hibás bázispárosodás következik be, azaz például egy adeninnel szemben guanin található, holott timinnek kellene lennie. Ekkor az excíziós repair vágja ki a hibás helyen lévő nukleotidot, majd pótolja egy odaillővel. Az repair rendszer a természetben a legkonzervatívabb rendszerek egyike, több esetében teljesen hasonlóan működik baktériumokban, gombákban és az emberben is. A repair rendszer hibájából adódóan a hibás DNS alapján hibás mRNS-lenyomat készülhet, majd arról funkcióképtelen vagy megváltozott funkciójú fehérje. Nem csoda, hogy olyan súlyos betegségeket alakít ki, mint a Bloom-szindróma vagy éppen a xeroderma pigmentosum.
Egyszerű DNS-repair (ez konkrétan excíziós repair)
Hasonlóan nagyon konzervatív mechanizmus biztosítja, hogy a már létrejött, de sérült sejtek eliminálják magukat, amire a természet a már emlegetett programozott sejthalál, azaz apoptózis formájában szolgáltatja a megoldást. A fokozott apoptotikus válasz gyakorlatilag mindig jelen van a neurodegeneratív betegségekben, míg az apoptotikus válasz hiánya tehető felelőssé egy halomnyi autoimmun betegség megjelenéséért.
Az előbb említetteken kívül a neopláziában, azaz a sejtek kóros felszaporodásában megfigyelték még azt is, hogy a telomeráz aktivitás (ez az enzim „varrja a kromoszómák sarkát” az osztódások után) a kelleténél nagyobb. A kromoszómák telomer régiója, ahogy a nevük is mutatja, a kromoszómák végein helyezkednek el, gyakorlatilag nem kódolnak semmilyen fehérjét, viszont minden sejtosztódás alkalmával rövidülnek, így egy-egy sejt osztódása a természetben korlátozott, tipikusan 30-50-re. Ha a kromoszóma vége „elkopik”, nem játszódik le teljes egészében több osztódás. Így válik érthetővé, hogy a telomeráz-rendszer helytelen működése hogyan eredményezi egy-egy sejt féktelen osztódását.
A posztban több esetben általánosságban írtam, ez viszont nem jelenti azt, hogy egy-egy mechanizmus hibája specifikus lenne egy-egy egy tumortípusra, viszont egy-egy enzimnek a hibája, ami egy-egy mechanizmus hibáját eredményezi, már tipikusan adott daganatos megbetegedés sajátossága.
Az egyszerűbb érthetőség kedvéért itt-ott szinonimaként használtam egy-egy gén nevét és az általa kódolt fehérje elnevezését, ezért mea culpa.
A ráktípusok és a ráktípusokra való hajlam örökletességére, a gyógyászatban használt potenciális terápiás targetekre később még valószínűleg visszatérek.
Gyakran találkozni azzal a véleménnyel, hogy programozni, na meg új informatikai technológiákat megtanulni könyvből nem lehet. Ami viszont legalább ilyen fontos: könyv nélkül normálisan pláne nem.
Nincs mese tehát, sokkal jobban járok, ha egy-egy témában inkább megrendelem a könyvet méregdrágán, hiszen olyan tudományos paradigmákról vagy technológiákról van szó, amivel kapcsolatban sehol nem találni átfogó és update művet a neten. Azaz például a molekuláris genetikán belül a rekombinációval kapcsolatban gyakorlatilag minden meghatározó kutatási eredmény összegereblyézhető a netről, ez olyan reménytelenül nagy energia- és időráfordítást igényelne attól, aki próbálkozna vele, hogy nyilván érdemesebb venni egy friss kötetet a témában, ami ugródeszkaként vagy kézikönyvként szolgálhat.
Web 2.0-ben rejlő technikai lehetőségek, ezek társadalmi hatásai, új értelmet nyerő public privacy, alkalmazott statisztika, sztochasztikus modellek, hibatűrő rendszerek, metamorf víruskódok, adatbányászat, adatbányászati módszerekkel kimutatható patternek olyan óriási rendszereken, mint a Facebook, Live!, Bebo vagy a MySpace… Csak néhány példát említettem, de a helyzet teljesen hasonló ahhoz, amit a második bekezdésben írtam: lehetetlen normálisan foglalkozni ezekkel, ha az ember fia nem boltolt be pár őt érdeklő témájú könyvet, amik nem ritkán „átússzák” az óceánt, mielőtt házhoz szállítaná a futár. Ha egy-egy könyvforgalmazó óriás keresőjében úgy adja meg a keresési feltételeket a kutató, hogy a csak a messzemenően specifikus témájú könyveket köpködje ki a kereső, még akkor is sokszor a bőség zavarával találja szembe magát.
Az elsősorban informatikai biztonsággal foglalkozó könyvet kiadására szakosodott No Starch Press webhelyére ugrottam fel a minap /*ez az a kiadó, amelyik korábban kiadta a világ legnagyobb hatású, legátfogóbb biztonságos programfejlesztéssel és bedugolással foglalkozó kötetét Hacking: The art of exploitation címmel*/. Ennél a kiadónál tudomásom szerint már csak a Syngress állt jobban rá az information security-privacy-témára, ahol gyakorlatilag csak ilyen témájú könyvek rendelhetők.
Visszatérve a No Starch Pressre, olyan kategóriába futottam bele ma, aminél még a processzorom ventillátora is megtorpant rövid ideig a meglepetés erejétől: szűk kört érdeklő tudományos témák mangásítva! Jól olvasod, mangásítva!
Én ugyan nem vagyok manga-fan, ismét az jutott eszembe, hogy megfelelő tálalással a mai világban mindent el lehet adni, népszerűsíteni, megszerettetni. Jelen esetben olyan tudományos témákat dolgoznak fel manga-könyvek formájában, amiktől normális ember agya egyébként is ledobná az ékszíjat. Csak néhány példa: adatbázis adminisztráció, statisztika, matematikai kalkulus manga formában.
Nemrég a Nature hozott le egy cikket arról, hogy egy vérszegénységben szenvedő fiút sikerült olyan génterápiában részesítenie egy francia és amerikai kutatókból álló csoportnak, aminek köszönhetően a beteg már rendszeres vértranszfúzió nélkül is teljes életet élhet. Miről is van szó valójában? A vérszegénységnek több típusa van, jelen esetben a fiú a béta-thalassaemia nevű örökletes betegségben szenvedett.
Mint tudjuk, a transzkripció folyamatában a DNS-ből mRNS „másolat” készül, ám maga a fehérje, ami a genetikai tulajdonságot kialakítja, nem közvetlenül ez alapján az mRNS alapján fog elkészülni, előtte az mRNS ún. RNS-érésen megy keresztül. Azaz többek közt a natív mRNS-kópiából még különböző enzimek kivagdossák azokat a szakaszokat, amik nem hordoznak semmilyen információt, ami a fehérje szintéziséhez szükséges lenne. Ezen kívül ekkor történik az ún. alternative splicing is, már ha történik, amiről érdemes néhány szót ejteni.
Ahogy azt mindenhol olvashattátok egy adott DNS-szakasz egy adott RNS-t kódol, ami egy adott makromolekula felépítéséhez szükséges információt tárol. Kivéve, amikor mégsem. (Személy szerint ezeket az eseteket szeretem a molekuláris biológiában.) Ugyanis az előbb emlegetett jelenség, azaz az alternative splicing folyamán az történik, hogy az mRNS-érés folyamán eltérő szakaszok vágódhatnak ki az eredeti RNS-ből, így értelemszerűen az érett RNS mást fog tartalmazni és többé-kevésbé más fehérjét is kódol.
A poszt elején emlegetett betegség molekuláris szintű oka pont az alternative splicing hiábjára vezethető vissza. Konkrétan arról van szó, hogy thalassaemia esetén egy DNS-szinten lévő mutáció következtében egy új hasítóhely található azon az mRNS-en, ami a vér oxigénjét szállító hemoglobin egyik láncát kódolja. Ilyen esetben az mRNS-érés folyamán történik egy olyan hasítás is, ahol nem kellene, aminek következményeként a normálistól eltérő szerkezetű hemoglobin molekula jön létre.
A jól ismert hemoglobin óriásmolekula egyébként 2 alfa és 2 béta alegységből áll, attól függően, hogy melyik mutációról van szó, kialakulhat alfa-thalassaemia vagy béta-thalassaemia is, ami annyit jelent, hogy az adott fehérje alegység instabil lesz, ami nem valami áldásos állapot egy létfontosságú fehérje esetében.
Sokáig a thalassaemiás betegekkel nem igen tudtak mit csinálni azon kívül, hogy folyamatosan friss vérrel látták el őket, a néhány nappal ezelőtti bejelentés jelentősége pont abban áll, hogy a thalassaemiás fiút olyan génterápiás módszert sikerült kezelni, aminek eredményeként már egyáltalán nem igényel rendszeres vérpótlást.
A génterápia igen tág fogalom, itt konkrétan arról van szó, hogy a kutatók a fiú csontvelőből származó őssejtjeinek DNS-ébe juttatták be a helyes hemoglobin szintézisért felelős gént. Az ilyet általában retrovirális vagy adenovirális vektorokkal oldják meg, ami annyit jelent, hogy az adott, bejuttatni kívánt gént kódoló DNS-szakaszt tartalmazó, de szaporodásra képtelen vírussal megfertőzik az adott őssejtet, azaz elérik, hogy a kívánt DNS-szakasz a sejtmagi DNS-be integrálódjon, majd az alapján ugyanúgy mRNS (majd fehérje) készüljön, mint bármely más sejtmagi génről. Amint ez megtörtént, a módosított génállománnyal rendelkező őssejteket visszajuttatják a betegbe, amelyiknek az immunrendszere jó esetben nem falja fel uzsonnára a nem kevés metodikai trükkel meghekkelt őssejteket, hanem az őssejtek utódsejtjei már a normális génváltozatokat fogják kódolni és kifejezni. Jelen esetben a hemoglobin stabil, normális béta-láncának termeléséről van szó a hibás géntermék termelése ellenében.
Az alábbi ábrán a génterápia kivitelezésének általános sémáját láthatjátok.
A különböző génterápiás módszerek megvalósításáról és ennek lehetőségeiről később még írok, amit viszont fontos figyelembe venni, hogy sok génterápiás próbálkozás egyszerűen azért vezetett kudarchoz, mert a módosított sejtek teljesen máshogy viselkedtek a sejttenyészetben, mint a beteg szervezetébe ültetve. Azzal kapcsolatban pedig sok esetben csak hipotézis született, hogy a módosított sejt miért viselkedett in vivo teljesen máshogy, mint ahogy az várható lett volna.
Az előző posztban tisztáztam a klónozás fogalmát, alternatív céljait és eloszlattam néhány ezzel kapcsolatos tévhitet, most pedig a szomatikus sejt-alapú klónozás technikai részleteiről írok röviden.
Ezzel a módszerrel az első, így legismertebb sikeresen klónozott emlős Dolly volt, akinek a tőgymirigy-sejteiből származott az a szövetminta, amiből a később beültetésre kerülő sejtmagot kinyerték. A kutatók tripszines emésztéssel elérték, hogy a szöveti sejtek egymástól elváljanak, majd az elválasztott sejteket táptalajra helyezték.
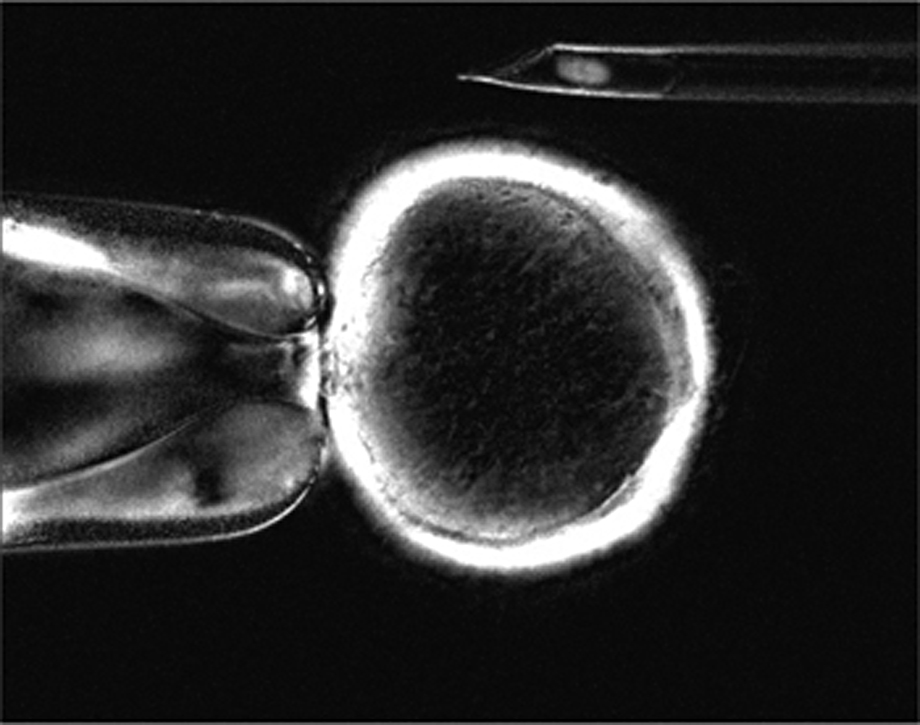
A táptalajra helyezett sejteket többszörös osztódásra, azaz proliferációra késztették, majd hirtelen megvonták a sejtek környezetében lévő tápfolyadékban az osztódásához szükséges komponenseket. Ennek következtében a sejtek sejtciklusuk azon pontján torpantak meg, amelyik stádiumban éppen voltak. A tenyészetből ezt követően azokat a nyugvó fázisban lévő, azaz nem osztódó sejteket választották ki, amik éppen az ún. G2 és M fázis közötti állapotban voltak. Minderre azért volt szükség, mert a sejtmagjától megfosztott recipiens petesejt szintén ilyen állapotban van, így tudja az idegen sejtmagot befogadni. A sejtmagjuktól megfosztott petesejteket ezidő alatt magnézium- és kálcium-ionoktól mentes pufferoldatban tartották. Amikor elérkezett a nagy pillanat, a petesejtet egy mikropipettával rögzítették, majd egy másik mikropipettával az előkészített sejtmagokból egyet a petesejt belsejébe juttattak.
írtam, hogy a petesejtet végig Mg- és Ca-mentes pufferben tartották, azt viszont nem, hogy erre a spontán bekövetkező Ca-szignál megjelenésének kiküszöbölése miatt volt szükség, ami a petesejtet alkalmatlanná tette volna a feladatra. Amikor viszont megtörtént a szomatikus sejt magjának beültetése, egy apró elektromos impulzus volt az, amelyik kalcium hiányában is kalciumszignált váltott ki a petesejtben, így a petesejt tényleges megtermékenyítés nélkül is osztódni kezdett.
A sikeresen osztódásnak indított petesejtből ezt követően néhány sejtes állapotú ébrény, ún. blasztocita jön létre, ezt ültették egy béranya-állat petevezetékébe, ahol elindult a tényleges embrionális fejlődés.
A siker mindenképp mérföldkőnek számít a biológia történetében, viszont az ismeretterjesztő írások is sokszor hajlamosak figyelmen kívül hagyni, hogy Dolly létrehozásához az akkori technikával 200-300 (!!) próbálkozásra volt szükség.
Persze Dolly születése után nem sokkal hozták létre Pollyt, azzal az óriási különbséggel, hogy Polly olyan klónozott emlősként látta meg a napvilágot, aminnek a klónozásához korábban kénkezelt sejtmagot használtak. Külön érdekesség, hogy Polly nem csak, hogy tartalmazza, de még expresszálni is képes volt a IX-es emberi véralvadási faktort, ami egyike azoknak a véralvadási faktoroknak, amik hiányában embernél a vérzékenység lép fel.
Klónozás. A téma annyira snassznak-kommersznek tűnik, hogy már-már cikivé válik írni róla ismeretterjesztő anyagot, olyan szempontból pedig nem vagyok egyszerű helyzetben, hogy ez egyáltalán nem területem. Mégis, a minap szóba került egy beszélgetés közben a klónozás témája és ismét szembesültem azzal a jelenséggel, amivel korábban találkoztam olyan témák kapcsán, amik eredetileg a molekuláris biológia és biotechnológia belügyeinek számítottak, majd átkerültek a közbeszédbe: sokszor totális a homály még a tájékozottabbak körében is.
A következő két posztban több aspektusból járom körül a klónozás témáját, viszont ésszerű terjedelmi korlátok miatt kerülöm a metodika részletes elemzését, ugyanakkor próbálom hangsúlyozni a sejtmagtranszferrel véghezvitt klónozás lényegét és ennek tudományos jelentőségét.
A klónozás* genetikailag teljesen ekvivalens élő szervezetek létrehozása mesterségesen, szomatikus, azaz érett testi sejt sejtmagának kiürített petesejtbe ültetésével. A szomatikus sejt beültetése után az alaposan meghekkelt petesejtet fejlődésnek indítják, ami elkezd osztódni, akárcsak a természetes módon megtermékenyített petesejtek. Természetesen több petesejten is elvégzik ugyanezt a műveletet, majd a néhány sejtes állapotban lévő embriók közül egyet aztán - emlős esetén - bejuttatják egy anyaállatba, ahol elindul az embrionális fejlődés, majd megszületik az utód. A megszülető utód genetikai szempontból tökéletesen egyezik azzal az egyeddel, amelyiknek egy testi sejtmagját a petesejtbe juttatták.
Ez az, amiről többé-kevésbé eddig is tudtunk, most pedig leírok pár dolgot, amivel megpróbálok eloszlatni néhány gyakori tévhitet, leírom a klónozás folyamatának leginkább lényegi lépéseit, így óhatatlan, hogy néhányatok egyik-másik megállapítást banálisnak fogja tartani, de bízom benne, hogy mindemellett sikerül több olvasónak minél több érdekes új információhoz jutnia a témában.
Gyakran teszik fel a kérdést, hogy ha klónoznának egy embert (ennek jogi vonatkozásai lentebb), akkor az ugyanúgy élne-e, ugyanolyan lenne-e a gondolkodásmódja, mint a donornak. Nos, mivel a donor és a klónja tökéletesen olyan, mintha egypetéjű ikrek lennének, ezzel meg is válaszoltam a kérdést: a környezettől is függ. Márpedig a megnyilvánuló tulajdonságok, az ún. fenotípus kialakításában az öröklött kifejeződő vagy látens genetikai tulajdonságokon túl a tág értelembe vett környezetnek is szerepe van, ami gyakorlatilag nem lehet teljesen azonos a két egyed esetén. Tehát például egy klónozott korábbi véres kezű diktátornak ugyan adott a hajlama arra, hogy olyan gondolkodásmódot és személyiségjegyeket mutasson, amint amilyen a néhai diktátoré volt, a biztosan eltérő kulturális- és neveltetési hatások miatt biztos, hogy nem ugyanolyan lenne az életvitele, nem ugyanazt az életutat járná be, ezt a témát hagyjuk is a science-fiction-szerzőkre. Az egyező génállomány miatt a teljesen azonos pszichológiai sajátosságok kialakulásának lehetősége viszont adott, így lehet, hogy lazán beérné egy klónozott Sztálin elvtárs azzal, hogy kitépkedi az elkapott legyek lábát egyenként, viszont véres kezű diktátor aligha válhatna belőle.
Néha hallok olyat, hogy a klónozás úgy en bloc tiltott, ami nem igaz. Országonként eltérő korlátozásokkal kísérleti állatok klónozhatók, tipikusan azért, mert őssejtek ilyen módon nyerhetők ki a leghatékonyabban, azaz még az embrionális fejlődés folyamán. Ilyen országonkénti eltérés például az, hogy egy klónozott kísérleti állatnál megengedett-e az, hogy kifejlődjön és megszülessen vagy csak néhány sejtes állapotig szabad fejlődni hagyni az embriót. A humán klónozás viszont szinte a világ összes országában más megítélés alá esik, ha úgy tetszik, más tészta, más szósszal. Ezzel kapcsolatban a magyar büntető törvénykönyv így rendelkezik: „Aki orvostudományi kutatás vagy beavatkozás során egymással genetikailag megegyező emberi egyedeket hoz létre, bűntettet követ el és öttől tíz évig terjedő szabadságvesztéssel büntethető.” (Btk. 173/G, p 1) Összefoglalva tehát klónozni éppenséggel lehet, de semmiképp sem reprodukciós célzattal.
A poszt második részében a klónozás technikai részleteivel foglalkozom, amit ide kattintva érhetsz el.
*a kötözködőknek írom, hogy igen, persze, a sejtmag-transzfer nélküli eljárás is klónozás, ezen kívül létezik a génklónozás, stb, ezek viszont nem tárgya a mostani két posztnak
Ma végülis nem volt erőm megszülni egy megfelelő minőségű cikket az olyan praenatalis diagnosztikai módszerek érdekességeiről, mint az amniocentesis vagy a chorion-boholy mintavétel, viszont kaptok helyette egy saját gyártmányú, háztáji bulvár bullshitet.
Valami miatt nem vált tömegfilmmé a Godsend, és nem lennék meglepve azon, ha ennek az lenne az oka, hogy a nézők egy részének egyszerűen nem esett le (spoiler a végén), hogy a filmben a doki mit is sumákolt el, amikor a házaspárnak klónozta a balesetben elvesztett fiukat.
Az az érdekes, hogy az egész egy átlagosan hülye amerikai thrillernek tűnik kb. a film kétharmad részéig és nekem teljesen az volt a benyomásom, hogy ismét egy olyan filmről van szó, amit azzal sikerült gallyra vágni, hogy egy nagyon nyomós tudományetikai kérdést okkultista bullshitettel raktak egy turmixgépbe a hollywoodi forgatókönyvírók. Aztán kiderül, hogy mégsem, viszont – mint írtam – szerintem nagyon sok nézőnek nem esik le.
Trailer itt, spoiler lentebb, ha gurítasz az egérrel.
Az egész sztori ötletgazdája sokkal jobban otthon lehetett a biotechnológiai módszerek humán alkalmazásának területén, mint ahogy azt sejteni lehetett volna: a film második felében az derül ki, hogy a doki nem csak klónozta a fiút, hanem egy másik, korábban meghalt szociopata kisfiú génjeit varrta bele a klónozott srác, Adam Ducan génállományába, amit persze a szülőknek nem mondott el. A DVD-s változatban egyébként több alternatív befejezés is látható. Ja, és természetesen még csak véletlenül se magyar szinkronnal nézd meg!
Eredetileg a http://karcsi.molbiolonline.com–ra írtam egy posztot a webhelyek látogatottságmérésével kapcsolatban, amit aztán gyorsan töröltem is. Itt pedig már nincs poszt két napja, most is galád módon egy informatikai témájú írással foglak büntetni benneteket.
Blogolni szép, blogolni jó, a blogolás megnyitja a szívcsakrákat. Valamelyik blogot csak az ismerősök olvassák jó esetben a haverok nyaggatása nélkül, érdeklődésből, ismét más blogoknak van egy nagyon szűk, viszont fix olvasótáboruk, amik néha napján az index.hu címoldalán robbannak és bizony, ki tudja, hogy a csillagoknak milyen szerencsés együttállása kell ahhoz, hogy egy blog abszolút sikerre vigye, azaz nagyon sok olvasót érjen el, már-már márkává nőjje ki magát.
Az elmúlt néhány napban bemutatott hypeok molbiolos fricskázása után kénytelenek lesztek beérni egy kis szájbarágóval. Igen. Algák, gyógyszermellékhatások és az evolúció. Mi köze van ezeknek egymáshoz?
Talán már középiskolában is volt róla szó, hogy a sejtek belsejében lévő sejtszervecskék, organellumok, úgy, mint a sejtek energiaháztartásáért felelős mitokondrium vagy éppen a növények esetében a fotoszintézisre képes színtestek részlegesen saját genetikai anyaggal, így programmal rendelkeznek, saját fehérjetermelő riboszómáik is vannak, ami persze nem jelenti azt, hogy önmagukban életképesek lennének.
Joggal vetődött fel a kérdés, hogy ezek a többi organellumhoz képest igencsak autonóm sejtszervecskék hogyan alakulhattak ki az evolúciós során. Erre a leginkább valószínű magyarázatot az endoszimbionta elmélet adta. Eszerint az elmélet szerint a mostani sejtmagvas, eukarióta sejtek ősei sejt-bekebelezéssel olyan bíborbaktériumokat és kékbaktériumokat faltak fel, amiket végül nem emésztettek meg, hanem ezek az ősi baktériumok együtt vígan élték világukat a mostani eukarioták ősei belsejében és ha olyan kedvük volt, önállóan osztódtak is. Az élővilágban a mai eukarióták közt a sejtszervecskéket felépítő fehérjék jókora része ugyan már átkerült a sejtmagi DNS-be, mégis számos olyan hasonlóságot mutatnak az önálló baktériumok életével molekuláris szinten, ami nem írható a véletlen számlájára.
Egy-egy újonnan tervezett gyógyszer-hatóanyag vizsgálatakor azonnal megnézik, hogy a sejtbe került vegyület hogyan hat a génműködésre, azaz létrehoz-e változást a DNS-ben, vagy más módon befolyásolja-e a sejtekben lévő gének kifejeződését. Ezek a vizsgálatok tehát elsősorban a sejtmagi örökítőanyagra és az abban lévő gének és géntermékek vizsgálatára terjednek ki. A felismerés, ami szerint gyógyszer-mellékhatásokért felelősek lehetnek a sejtmagon kívüli, ún. extranukleáris öröklődésben szerepet játszó elemek is, mindössze néhány évtizedes.
Megfigyelték, hogy a sejtek energiaszolgáltató szervecskéiben, a mitokondriumokban a transzkripció folyamatát, azaz azt a lépést, aminek során a DNS alapján RNS-kópia készül, a rifampicin nevű antibiotikum blokkolja, így a mitokondrium értelemszerűen nem lesz képes a számára szükséges fehérje előállítására. Később a tetraciklinről, majd a klóramfenikolról bizonyosodott be, hogy a baktériumok elpusztítása mellett szintén gallyra vágják a miktokondriumok fehérjeszintézisét. Ebben utóbbi két vegyület a transzlációt, azaz az RNS-minta alapján történő fehérjeszintézist blokkolja le, miközben a sejt többi részében a fehérjeszintézisre egyáltalán nincs hatással. A magyarázat pedig az, hogy a mitokondriumokban nemcsak a saját DNS szerkezete mutat óriási hasonlóságokat a baktériumok DNS-ével, hanem saját transzkripciós és transzlációs enzimjeik is nagyon hasonlítanak vagy éppen azonosak azokkal a transzkripciós és transzlációs folyamatokban működő enzimekkel, amik baktériumokban vannak jelen.
A sejtorganellumok működésével kapcsolatos részleteket ma már az extranukleáris öröklődéssel foglalkozó kutatók részletekbe menően ismerik, ami a gyakorlatban annyit jelent, hogy az egykor elméleti jelentőségűnek tűnt felismeréseket ma már minden gyógyszerfejlesztésnél figyelembe veszik.
Ha olyan perspektívából nézem a dolgot, hogy hogyan is függ össze a betegekben felszaporodó baktériumokat elpusztító antibiotikum és maga a mellékhatás, az egyik legfontosabb, ami figyelembe kell venni, hogy a létfontosságú mitokondriumok száma sejttípusonként nagyon eltérő, függően az illető sejt, értelemszerűen az adott sejtekből épülő szövet energiaszükségletétől. így molekuláris erőműveink átmeneti károsodása teljesen máshogy fog hatni olyan szervekre, amiknél az energiaáramlás csekély, mint olyan szervek esetén, ahol intenzív. Egyese sejtjeinket egyetlen mitokondrium is kiszolgálhatja, míg megint más sejtekben több ezerre van szükség belőlük.
Természetesen nem minden antibiotikum hat a mitokondriumok működésére, viszont az egyértelműen kimutatható, hogy adott antibiotikum adott baktériumtörzs egyedeinek elpusztítása mellett mennyire nyírja a mitokondriumokat is, így az ebből adódó mellékhatás még vállalható-e klinikai szinten.
A mitokondirális öröklődésről, pláne az antibiotikumok mitokonriumokra kifejtett hatásáról közérthető videót természetesen nem találtam, olyat viszont igen, ami érthetően bemutatja, a mitokondrium energiatermelő funkcióját.
Mint sok más, globális egészségügyi probléma területén, az AIDS gyógyításával kapcsolatban is rendre röppennek fel hírek azzal kapcsolatban, hogy megtalálták a betegség ellenszerét.
Kezdeném azzal, hogy maga a kialakult betegség, az AIDS a tünetegyüttest jelenti, ami a fertőzés egy viszonylag késői stádiumában jelenik meg, míg maga a HIV-fertőzés annyit jelent, hogy valakinek a keringési rendszerében már ott van a vírus, de nem feltétlenül beteg.
Az AIDS Research and Therapy egy augusztusi cikke szerint izraeli kutatóknak ezúttal sikerült olyan hatóanyagot előállítaniuk, amik az eddigieknél sokkal inkább célzott módon gátolják a vírus terjedését, mégpedig úgy, hogy csak a fertőzött sejteket pusztítják el, méghozzá korábban, mint hasonló hatásmechnizmussal működő társaik.
A HIV-vírus az ún. retrovírusok közé tartozik, fertőzésének módja részletekbe menően ismert, ez tette lehetővé olyan, teljesen eltérő támadáspontú hatóanyagok kidolgozását eddig is, amiket egymással kombinálva évtizedekig késleltethető a fertőzöttekben magának a betegségnek, a teljes immundeficitnek a kialakulása. A HIV-fertőzés során a vírus jellemzően CD4-es és CD8-as T-limfociták, azaz fehérvérsejtek receptorain, azaz kötőfehérjéin tapadnak meg, jutnak be a sejtbe, majd saját RNS-üket a szépnevű reverz transzkriptáz enzim segítségével több lépésben a gazdasejt DNS-ébe másolják. A megváltozott genetikai programmal rendelkező fehérvérsejtek innentől kezdik el gyártani a vírus új példányait, aminek egyik hatása, hogy a sejt sok-sok apró víruspartikulum legyártása után a sejt elpusztul, a vírusok a keringésbe jutva más sejteket fertőzhetnek meg. Magának a betegségnek az oka pedig az, hogy pont olyan sejteket használ a vírus önmaga felszaporításához, így elpusztításához, amik kulcs fontosságúak az immunrendszer szabályozásában.
Terveztem, hogy később részletesen is írok egy-egy vírusfertőzés molekuláris szintű mechanizmusairól, most csak nagyon röviden néhányat említek, amiknek a megismerése a klinikumban is használható hatóanyagok készítését tette lehetővé. Az egyik támadáspont a reverz transzkripció folyamatának gátlása, azaz annak a mechnizmusnak a megakadályozása, hogy a vírus RNS-e a gazdasejt DNS-ébe íródjon át, így gyakorlatilag átprogramozva a sejtet, ahogy azt korábban is írtam. A másik fő támadáspont a vírus-specifikus proteáz enzimet gátolja. Ez az enzim, ami azt eredményezi, hogy a módosított DNS alapján készülő vírusfehérjék megfelelő módon szeletelődjenek azért, hogy azok később a vírus építőelemei lehessenek. Mindkét módszernek számos altípusa van, de gyakorlatilag a HIV az összes ellen képes volt eddig rezisztenciát kifejleszteni.
A posztom elejében szóba hozott felfedezés jelentősége abban áll, hogy a kutatók olyan proteáz inhibitorokkal álltak elő, amik a jelenleginél – a cikk szerint – hatékonyabbak, hiszen sokkal előbb ölik meg a fertőzött fehérvérsejteket, mielőtt azok halomszámra gyártanák az új vírusokat.
Tehát ismétlem, arról szó sincs, hogy bulvárosan szólva „feltalálták volna a HIV-ellenszerét”, csak arról, hogy találtak egy új, eddigieknél hatékonyabb proteáz inhibitort, ami ellen ha a vírus nem fejleszt ki rezisztenciát, összességében azt eredményezi, hogy a vírusok csak minimális mértékben képesek önmagukat felszaporíttatni az immunrendszerben (hiszen a vírusgyárakká vált fehérvérsejtek idejekorán elpusztulnak), így az AIDS kialakulása még inkább elodázható.
Bizony a molekuláris genetikai folyamatok közt is találunk olyat, amire metaforaként azt a legtalálóbb mondani: a fagyi visszanyal.
Jól ismert, hogy a különböző hatásmechanizmusú antibiotikumok elterjedésével a baktériumok ezekre a hatóanyagokra egyre inkább ellenállóvá, rezisztenssé váltak. Sőt, több helyen lehetett már olvasni ún. multidrog-rezisztenciát mutató szuperbaktériumokról, amiket az sem zavartatja, ha az egykoron legelvetemültebbnek számító antibiotikum-kombókkal találják szembe magukat.
Ami viszont már jóval kevésbé köztudott, hogy a baktériumok milyen molekuláris változásokon is mennek keresztül több nemzedéken keresztül ahhoz, hogy csemetéik túlélőbbek legyenek elődeiknél.
Ahány féle antibiotikum-csoport, annyiféle hatásmechanizmus, mindnek a bemutatása jóval meghaladná egy blogposzt kereteit, így most csak az egyikre térek ki, a legismertebb antibiotikum ellen kialakuló rezisztenciáról írok.
A legkorábban felfedezett, egyes irodalmi források szerint csak a második világháború táján több, mint 150-160 tonna (!!) mennyiségben legyártott pencillin és a most használt penicillin kifejezés alatt valójában egy igen népes vegyületcsoportot értünk, ezen vegyületek közös tulajdonsága a szerkezetükben megtalálható amino-penicillánsav csoport, aminek a tényleges antibakteriális hatás tulajdonítható.
A penicillin hatásmechanizmusát tekintve az ún. béta-laktám antibiotikumok közé tartozik.
Miután egy béta-laktám típusú antibiotikum a baktérium belsejébe jutott, megakadályozza a baktérium azon enzimjeinek működését, amik bizonyos baktériumok döntően peptidoglikán anyagú sejtfalának kialakításához szükségesek. Így a folyamat eredményeként a baktérium sejtfala nem tud szabályosan kialakulni, aminek következtében a baktérium bánatában egy kapcsolt folyamat során autolizineket, azaz önmagát a baktériumot felemésztő enzimeket aktivál, amivel be is következik a molekuláris szintű baktérium-kamikaze. Értelemszerűen a penicillin, ahogy a béta-laktám támadáspontú hatóanyagok többi tagja is, csak olyan baktériumok esetén hatásos, amiknek a patogenitása és immunrendszerrel szemben való ellenálló képessége a peptidoglikán sejtfaltól is függ.
A baktériumok viszont néhány évtizednyi, mondhatni „mikroevolúciós” folyamat során olyan mutációkon mentek keresztül, amik hatására bizonyos génjeik képesek a béta-laktám-típusú antibiotikumokat inaktiválni. Ilyen baktérium-géneket kimutattak baktérium-kromoszómákban illetve a baktériumban a teljes baktérium osztódása nélkül, a baktérium belsejében önmagukban felszaporodni képes ún. plazmidokban, amik apró, gyűrű alakba zárt DNS-részletek és nem feltétlenül öröklődnek át egyik baktérium generációról a másikra.
Miről is van szó, miben áll a rezisztencia kialakulásának lényege molekuláris szinten?
(1) a penicilinek elterjedésével a baktériumok evolúciójuk során versenyt futottak az antibiotikumokkal, csak éppen gyorsabban, mint ahogy azt korábban sejteni lehetett volna. Így például olyan gének jelentek meg bennük, amik a pencillinek tényleges hatásáért felelős amino-penicillánsav egyik gyűrűjét felvágják, így inaktiválják a hatóanyagot. A evolúció persze szorgosabb volt ezen a téren, mint ahogy azt bárki gondolná: már jóval több, mint 200 gént azonosítottak, amik kifejeződésekor béta-laktamáz aktivitású enzim jelenik meg. És ez még csak az egyik típusú mechanizmus egy adott hatóanyag típusra!
(2) Ismét más, a baktériumokban újonnan megjelenő gének olyan enzimeket kódolnak, amik az amino-penicillánsav gyűrűt nem hasítják el ugyan, viszont megfogják a teljes antibiotikumot, majd olyan konformáció változást /*szerkezetbeli változást*/ hoznak bennük létre, amik ilyen módon teszik lehetetlenné az antibiotikum működését.
(3) A poszt első felében azzal kezdtem, hogy az antibiotikumoknak a baktériumsejtek belsejébe kell jutniuk azért, hogy hatásukat kifejthessék. Hát persze, hogy ezt a trükköt is kihasználja a baktériumok evolúciója olyan módon, hogy megváltoztatják a baktériumok sejtfalának antibiotikummal szembeni áteresztő képességét, ún. permeabilitását.
A pencililin széles körben eredetileg Staphyllococcus törzsek ellen vetették be. Ennek röpke néhány évtizedes evolúciós vonatkozású eredménye az lett, hogy a egy-egy betegben elszigetelten kialakul fertőzéseknél ezen törzsek rezisztenciája 80-90%-os, míg kórházakban előforduló baktériumokban 95-99%-os. (Az utóbbi adatot egyik könyvemből puskáztam ki, azóta már változhatott a helyzet, de a lényeg ugyanaz. )
Későbbi posztokban más kórokozók, más támadáspontú vegyületek és más molekuláris szintű védekezési mechanizmusok megjelenéséről is beszámolok majd.
Nem tudom eldönteni, hogy itt valaki most nagyon nagyot akart villantani és ennek érdekében a kutatás valamelyik pontján egyszerűen "meghekkelte a rendszert" (l. alább), esetleg nagyobb szenzációként lett kikommunikálva a tömegsajtóba a hír, ha igaz, differenciálódott, érett bőrsejtekből hoztak létre májsejteket.
Ismét egy kis szájbarágóval indítanék: testünket több száz különböző típusú szövet építi fel, amik a szöveti differenciálódás folyamatában válnak tényleges szöveti sejtekké még az embrionális korban őssejtekből, amik multipotens/totipotens/pluripotens állapotban lehetnek, azaz potenciálisan bármilyen szövet fejlődhet ki belőlük. Az őssejtekkel kapcsolatos kutatásoknál az embrionális őssejtek jelentősége abban mutatkozik meg, hogy nem kell érett szöveti környezetben vájkálni (pl. csontvelőben) ahhoz, hogy a kutatók őssejtekhez jussanak. A lényeg, hogy ha egy sejt valamilyen fejlődési pályára ráállt és elkezdett fejlődni egy bizonyos szövettípusú sejtjévé, a folyamat nem fordulhat vissza, soha nem lehet belőle ismét őssejt. Ez alól kivételeket csak a növényvilágban találunk (pl. gyökérmerisztéma-sejtek, ha jól emlékszem).
Ehhez képest, aki követi az őssejtekkel kapcsolatos irodalmi adatokat, észreveheti, hogy szinte slágerként, rendszeresen előáll valaki azzal, hogy debizonymégis… Újabban a ScienceDialy a Journal of Clinical Investigation legfrissebb számából vette át azt a cikket, amiből kiderül, hogy a Cambridge Egyetem kutatói bőrsejtekből előállított májsejtek.
A hírügynökségek persze már bőrsejtek "átprogramozásáról" írtak, ami még inkább bulvár szagúvá teszi az egészet. Az SicenceDialy számomra kicsit zavaros cikke hivatkozva persze nem felejti el megjegyezni, hogy mekkora jelentősége is lehet különböző betegségek gyógyításában annak, hogy májsejteket állítottak elő bőr sejtjeiből. Mintha embrionális őssejtekből nem lehetne (ráadásul jóval egyszerűbben). A cikk valóságtartalmát egyébként nem vitatom, nem kompetenciám, viszont aki őssejt-ügyileg kevésbé tájékozott, az jó, ha plusz információként még ezt-azt szem előtt tart, ami alapján már mondhatjuk, hogy a mostani bejelentés tényleg kicsit túl van hype-olva. Ezekből a plusz infókból gyűjtöttem egy csokorral.
(1) Nem derül ki, hogy pontosan milyen metodikákat használtak a kísérlet folyamán, így – mint minden más helyen – felvetődik a módszertani és az egyszerű mérési hiba lehetősége. Az utóbbi pár évben kiderül, hogy olyan szöveti környezetben is előfordulnak őssejtek, ahol nem gondolták volna (hogy szöveti sérülés esetén azok gyorsan osztódva minél gyorsabban ki tudják pótolni a sérült szövetet egészségessel). Így nem csoda, ha többünkben felvetődik az, hogy esetleg pont olyan sejteket pecáztak ki a bőrt alkotó hámszöveti környezetből, amik azért mégsem voltak teljesen differenciálódott sejtek. Ezek pedig megfelelő szignálok és növekedési faktorok hatására fejlődhettek májsejtekké. Ez esetben viszont szó sincs róla, hogy differenciálódott sejtek differenciálódtak volna át valami mássá.
(2) Érdekesnek ugyan érdekes, kérdéses, hogy a tudomány jelen állása szerint mennyi értelme van egy olyan módszernek, aminek során differenciálódott sejtekből állítanak elő más differenciálódott sejteket, hiszen ez ma már rutinszerű kezdeti fázisban lévő embriók sejtjeinek vagy például a köldökzsinór vérében található őssejtek felhasználásával. Ez viszont önmagában persze még nem csökkenti az új tudományos felismerés értékét.
(3) Az a megállapítás, hogy „Ez a technológia kikerüli a humán embriók használatának szükségességét”, első olvasásra több, mint manipulatív. Ugyanis őssejtkutatásokban nem feltétlenül kell humán embriókat felhasználni (sőt, az a ritkább), ott van helyette kismillió faj, aminek az embriói felhasználhatóak, aztán ha kimondottan egy humán-vonatkozású őssejt-kísérlet adódik, akkor kell csak humán őssejteket használni. Másrészt az embrionális őssejtekkel kapcsolatos bioetikai vita egyre inkább lejárt lemeznek tűnik, a jelenség kísértetiesen hasonlít a néhány évtizeddel ezelőtti GMO-vitára, ami szintén a molekuláris biológia eddigi rövid történetének egyik legnagyobb, ha nem a legnagyobb hypeja volt.
Roppant szar szóviccel élve – nem tudom, hogy hány bőrt lehet még lehúzni az őssejt témáról, de itt is megfigyelhető az, ami a tudományos kutatásoknál általános: elviheti a figyelmet a fontosabb alapkutatás eredményeiről, ez persze nem a kutatók hibája.
Az a blog, ahonnan a fotót átvettem, azért merészebben fogalmazott a témában közel három évvel ezelőtt. Akit érdekel:
Növények. A magasabb rendűek közt gyakorlatilag nincs, amelyik képes lenne megváltoztatni a helyét, ezért az evolúcióban kénytelenek voltak menekülés helyett más védekezési stratégiákat kialakítani ellenségeik, a növényevő állatok ellen.
Pont ez az oka a növényvilágban előforduló, különböző vegyületcsoportokba tartozó mérgek rendkívüli változatosságának is – a rovar megrágja, majd elpusztul, a növény pedig megmenekül.
A bevezetőben valószínűleg sokak számára nyílt kapukat döngettem, így most rá is térek a lényegre. Nemrég több helyen is lehetett olvasni, az egyik legtrükkösebb növényi védekezési stratégiáról. A Planck Intézet kutatóinak sikerült igazolni, hogy a dohányszender hernyójának nyála a megrágott dohánynövény néhány sérüléskor felszabaduló illékony vegyületét úgy változtatja meg, hogy annak szaga pont dohányszender-hernyókkal is táplálkozó, ragadozó poloskákat vonza oda.
Hogyan is kapcsolódik mindez a molekuláris biológiához? Az összefüggés tisztázása lehetetlen lett volna anélkül, hogy kimutassák: a megrágott növényből felszabaduló illékony GLV szerkezetét az egyik izomeráz enzim alakítja át a hernyó nyálának hatására olyan illatú anyaggá, amit a poloska ténylegesen megérez.
A folyamatot később kísérletileg is igazolták. Összességében tehát a dohány is megmarad a poloska is jóllakik.
A D-vitamin jelentősége jól ismert. Az viszont jórészt ismeretlen volt, hogy molekuláris szinten hogyan hat a D-vitamin, hogyan is befolyásolja az élettani folyamatokat. Körülbelül mostanáig.
Nemrég jelent meg egy cikk a Genome Researchben, amiben egy nemzetközi staff kutatói arról számolnak be, hogy a D-vitamin jelenléte hogyan hat bizonyos gének kifejeződésére. Érdekesség, hogy az anyagot, amit valószínűleg az MTI gányolt először képernyőre magyarul, elképesztően sok lap átvett úgy, hogy a magyar nyelvű cikkek az átvett formájukban gyakorlatilag értelmetlenek. Ködösen írnak valami receptorról, meg DNS-kötésről, de hogy mi, mivel, hogyan, na az teljesen kimaradt. Ezért is gondoltam rá, hogy leírom vázlatosan, de logikusan itt, hogy miről is van szó.
Mint tudjuk, a sejtmag DNS-ében kódolt kb. 30100 gén hol működik, hol nem, attól függően, hogy van-e rá szükség vagy éppen nincs. A gén kifejeződésének, expressziójának nevezem azt, amikor a DNS adott gént kódoló szakasza szabaddá válik, majd arról mRNS-kópia (szintén fonálszerű óriásmolekula, ami már jórészt csak magának a génnek a tervrajzát hordozza) készül, ami a sejtmagból kikerül. Miután kikerült, a riboszómáknak nevezett sejtszervecskék és tRNS-ek az mRNS-kópia alapján elkészítik magát a fehérjét, ami a sejtből vagy kijut vagy nem, lényeg, hogy a fehérje, mint géntermék felelős magáért a genetikai tulajdonságért. Az pedig, hogy a magban lévő DNS-nek melyik része szabad, azaz átírható éppen-e, DNS-kötő fehérjék határozzák meg, amik ún. magmembrán fehérjékkel vannak kapcsolatban. Tehát bizonyos magmembrán-fehérjék szabályozzák magát a génexpressziót. Ennyi lett volna a bevezető szájbarágó, most rátérek, hogy hogyan kapcsolódik mindez a D-vitaminhoz.
Amit a hírportálok amolyan sejtelmesen VDR-nek becéznek anélkül, hogy leírnák pontosan, hogy az mi is, nem más, mint az ún. sejtmagi „vitamin D receptor”. A kutatók azt találták, hogy ehhez a receptorhoz kötődve pontosan 229 gén expressziójának szabályozásában játszik be a D-vitamin, amik közt több olyan is van, amit korábban már összefüggésbe hoztak bizonyos betegségek megjelenésével.
A D-vitamin amint a saját magi membránreceptorához kötődik (VDR), olyan szignált vált ki, aminek hatására a sejtmag belsejében a DNS adott, géneket hordozó szakaszai szabaddá válnak, ami lehetővé teszi, hogy az érintett gének kifejeződjenek. Tovább bonyolítja a dolgot, hogy a D-vitamin pleiotrop, azaz nagyon sok génre való hatása miatt olyan gének is érintettek, amik ún. transzkripciós faktorként működnek. Ezek olyan gének, amik semmi más sem csinálnak (ebben mondjuk soha senki sem lehet biztos), csak más gének átíródását szabályozzák.
Az új felfedezés kiemeli, hogy a szóban forgó mechanizmus olyan génekre hat, amik összefüggésbe hozhatók a Crohn-betegséggel, a demenciával, az izületi gyulladásokkal, adott immunrendszeri folyamatokkal, amik szerepet játszanak a scelrosis multiplex kialakulásában és jónéhány tumortípusnál.
Mit jelent ez a gyakorlatban? Sokat, de az ilyen típusú pleiotrop-jelleg nem olyan ritka a valóságban, egy-egy egészséges vagy kóros élettani folyamat lefolyása pedig mindig egyszerre nagyon sok gén működésétől (expressziós mintázat) függ.
Addig is zabáljuk sok D-vitamint, mert az jó, de az előbbiből adódóan nem csodaszer.
Amerika egy perfetisiszta ország, ezt tudjuk jól. Valóságos kultúrája van annak, hogy mindenki perel mindenkit minden szarért. Igazából nem is az a legrémisztőbb, hogy egyesek azért perelnek, hogy megmogyorózzák a konkurenciát, kártérítést csikarjanak ki vagy hasonló financiális érdek vezérli őket. A legijesztőbbek a meggyőződéses hülyék, akik mindenféle szakmaiságot nélkülöző érvekkel hozakodtak elő egy per folyamán, mégis nekik ad igazat a bíróság.
Ahogy azt a Wall Street Journal egy nem túl régi cikkében lehet olvasni, a bíróság a zöldek kezdeményezésére egyszerűen betiltotta a genetikailag módosított cukorrépát, ami az ottani cukoripar alapja. Az érvelés elképesztően ostoba: nem előzte meg a GM cukorrépa bevezetését hatástanulmány.
Miről is van szó? A zöldek kultikus ősellensége, a Monsanto olyan genetikailag módosított cukorrépát hozott létre, majd juttatott a gazdákhoz, aminek hatására a répa rezisztens a glifoszfát-alapú gyomirtóra, így a parcellákra kijuttatott gyomirtó alkalmazása során a gyom elpusztul, a termény viszont ép marad. Egy szóval egyszerűbb és hatékonyabb a termesztés, hiszen nincs szükség arra, hogy a gyomirtó alkalmazása nélkül fenntartott cukorrépa-parcellákon folyamatosan kikapálják a megjelenő gyomot, hiszen nem-GM répa esetén ez a gyomirtó nem alkalmazható. Egyszerűbb, így olcsóbb is.
Az érvelések gyakorlatilag a témában már teljesen megszokott szűklátókörű sületlenségek. Kifogásként hozták fel, hogy a GM répa esetleg hibridizálódik a nem-GM-répával. WTF? Mintha a természetben nem jönnének létre egyébként is nap, mint nap növényi hibridek a legkülönbözőbb körülmények és sokszor egymáshoz nem is olyan közeli fajok között. A másik érv szintén amolyan örök klasszikusként zendült fel: fennáll a veszélye annak, hogy a gyomirtószerrel szemben rezisztens gyomok jelennek majd meg, aminek az ostobaságát ahhoz tudnám hasonlítani, mintha egy súlyos tüdőgyulladásban szenvedő betegtől megvonnák az antibiotikumot azért, hogy a benne lévő pneumonae-k nehogy rezisztenssé váljanak és kikerüljenek a betegből, aztán letámadják az emberiséget.
Nem is taglalom tovább most ezt a GMO pro és kontra vitát, hiszen ezt már valójában jóideje eldöntötte a tudomány, majd ezt követően a piaci szereplők. Ami miatt ezt most mégis megírtam, az annak az ijesztő ténye, hogy az amerikai bíróságon ezek szerint tényleg nincs akkora debilség, amit ne lehetne keresztül vinni, még abban az esetben is, ha az társadalmi szempontból kimondottan káros. Hiszen ha visszatérnek a hagyományos cukorrépához és a hagyományos növényvédelmi módszerekhez, az nyilván alaposan megdobja majd a cukor piaci árát. Nem csak az USA-ban, hanem mindenhol.
Érdekes módon, ha valakinek van egy vadalma fa a kertjében, a szomszéd pedig ültet valamilyen nemesített almát, normális ember nem rohan a bíróságra azzal, hogy a vadalmafájával a szomszéd nemesített almafája majd hibridet képezhet. Nem, ha több hektárnyi, különböző fajtájú almát termesztenek egymás melletti parcellákon, ez akkor sem jut eszébe senkinek, mint különös kockázat. Pedig lényegi különbség nincs biológiai szempontból. Az óriási különbség az, hogy például a Jonagold vagy a Summered almafajtákat emlékeim szerint klasszikus nemesítési módszerekkel hozták létre sok-sok generáción keresztül, nem pedig közvetlenül gének módosításával, ahogy az néhány évtizeddel ezelőtt lehetővé vált. Ennyi.
Az utóbbit viszont minden szalonhülyének elleneznie kell. Hogy miért? Izé, leginkább, mert csak. Természetesen a direkt genetikai módosításnak is megvan az egészségügyi, ökológiai kockázata, akkor, ha egyszerűen rosszul csinálják. Viszont elcseszni mindent lehet.
Nemrég a PLoS Biologyn közölték le, hogy a trippert okozó baktérium fertőzési stratégiája trükkösebb, mint ahogyan azt korábban sejteni lehetett volna.
A húgyivari szervek gyulladását okozó Neisseria gonorrhoeae baktérium fehérjefonalaival a gazdasejt membránjához horgonyozza magát, ezt követően pedig a saját felszínét úgy változtatja meg, hogy az alapján az immunrendszer ne ismerje fel. (Mint ismeretes, az immunrendszer erre specializálódott sejtjei a parazitákat és a tumorsejteket sejtfelszíni fehérjéik alapján tudják azonosítani, majd az ennek megfelelő immunreakciót beindítani.) Amint ez sikerült, gyakorlatilag belemászik, majd felszaporodik a sejtekben, ami molekuláris szinten elindítja a betegség tényleges folyamatát.
A felfedezés jelentőségét abban lehet, hogy a horgonyzó molekulák működésének jobb megismerése hozzájárulhat a kimondottan ezt célzó antibiotikumok vagy más, a horgonyzás mechanizmusát megfékezni próbáló hatóanyagok fejlesztésében.


 Nemrég a
Nemrég a  A személyre szabott genetika/genomika témában természetesen
A személyre szabott genetika/genomika témában természetesen  Érkezik, amit legalább hónapok óta szinte biztosra lehetett tudni – a
Érkezik, amit legalább hónapok óta szinte biztosra lehetett tudni – a 
 Az érdeklődő emberben biztosan felmerült már az a kérdés, hogy mennyire is bonyolult egy élő sejt működése? Röviden megválaszolva: nagyon. Azok az anyagcsereutak, amik leírják, hogy milyen anyag milyen más anyaggá alakul át egy-egy enzim hatására, nem meglepő módon hálózatot alkotnak. Jópár enzim (és annak termékei) közül némelyiknek a defektusa vagy hiánya teljesen összeegyeztethetetlen az élettel, megint más esetben sokáig azt hitték, hogy összeegyeztethetetlen az élettel, aztán kiderül, hogy mégsem. Itt konkrétan
Az érdeklődő emberben biztosan felmerült már az a kérdés, hogy mennyire is bonyolult egy élő sejt működése? Röviden megválaszolva: nagyon. Azok az anyagcsereutak, amik leírják, hogy milyen anyag milyen más anyaggá alakul át egy-egy enzim hatására, nem meglepő módon hálózatot alkotnak. Jópár enzim (és annak termékei) közül némelyiknek a defektusa vagy hiánya teljesen összeegyeztethetetlen az élettel, megint más esetben sokáig azt hitték, hogy összeegyeztethetetlen az élettel, aztán kiderül, hogy mégsem. Itt konkrétan 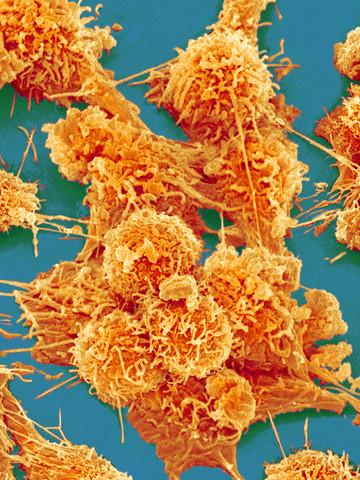 Már napokkal ezelőtt elhatároztam, hogy dobok egy posztot korunk egyik népbetegségéről, a daganatos megbetegedésekről. Onkológus ugyan nem vagyok, az már előre látszott, hogy nem leszek könnyű helyzetben, ha erről olyan posztot szeretnék írni, amiben a tumoros megbetegedések alapvonásait szakszerűen, mégis könnyen elfogadható nyelvezettel próbálom tálalni.
Már napokkal ezelőtt elhatároztam, hogy dobok egy posztot korunk egyik népbetegségéről, a daganatos megbetegedésekről. Onkológus ugyan nem vagyok, az már előre látszott, hogy nem leszek könnyű helyzetben, ha erről olyan posztot szeretnék írni, amiben a tumoros megbetegedések alapvonásait szakszerűen, mégis könnyen elfogadható nyelvezettel próbálom tálalni.  Gyakran találkozni azzal a véleménnyel, hogy programozni, na meg új informatikai technológiákat megtanulni könyvből nem lehet. Ami viszont legalább ilyen fontos: könyv nélkül normálisan pláne nem.
Gyakran találkozni azzal a véleménnyel, hogy programozni, na meg új informatikai technológiákat megtanulni könyvből nem lehet. Ami viszont legalább ilyen fontos: könyv nélkül normálisan pláne nem.  Nemrég a Nature hozott le egy cikket arról, hogy egy vérszegénységben szenvedő fiút sikerült olyan génterápiában részesítenie egy francia és amerikai kutatókból álló csoportnak, aminek köszönhetően a beteg már rendszeres vértranszfúzió nélkül is teljes életet élhet.
Nemrég a Nature hozott le egy cikket arról, hogy egy vérszegénységben szenvedő fiút sikerült olyan génterápiában részesítenie egy francia és amerikai kutatókból álló csoportnak, aminek köszönhetően a beteg már rendszeres vértranszfúzió nélkül is teljes életet élhet. 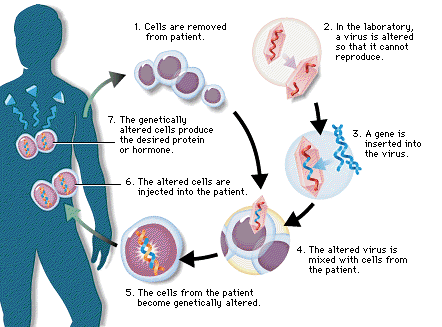
 A sejtmagjuktól megfosztott petesejteket ezidő alatt magnézium- és kálcium-ionoktól mentes pufferoldatban tartották. Amikor elérkezett a nagy pillanat, a petesejtet egy mikropipettával rögzítették, majd egy másik mikropipettával az előkészített sejtmagokból egyet a petesejt belsejébe juttattak.
A sejtmagjuktól megfosztott petesejteket ezidő alatt magnézium- és kálcium-ionoktól mentes pufferoldatban tartották. Amikor elérkezett a nagy pillanat, a petesejtet egy mikropipettával rögzítették, majd egy másik mikropipettával az előkészített sejtmagokból egyet a petesejt belsejébe juttattak.  írtam, hogy a petesejtet végig Mg- és Ca-mentes pufferben tartották, azt viszont nem, hogy erre a spontán bekövetkező Ca-szignál megjelenésének kiküszöbölése miatt volt szükség, ami a petesejtet alkalmatlanná tette volna a feladatra. Amikor viszont megtörtént a szomatikus sejt magjának beültetése, egy apró elektromos impulzus volt az, amelyik kalcium hiányában is kalciumszignált váltott ki a petesejtben, így a petesejt tényleges megtermékenyítés nélkül is osztódni kezdett.
írtam, hogy a petesejtet végig Mg- és Ca-mentes pufferben tartották, azt viszont nem, hogy erre a spontán bekövetkező Ca-szignál megjelenésének kiküszöbölése miatt volt szükség, ami a petesejtet alkalmatlanná tette volna a feladatra. Amikor viszont megtörtént a szomatikus sejt magjának beültetése, egy apró elektromos impulzus volt az, amelyik kalcium hiányában is kalciumszignált váltott ki a petesejtben, így a petesejt tényleges megtermékenyítés nélkül is osztódni kezdett.  Klónozás. A téma annyira snassznak-kommersznek tűnik, hogy már-már cikivé válik írni róla ismeretterjesztő anyagot, olyan szempontból pedig nem vagyok egyszerű helyzetben, hogy ez egyáltalán nem területem. Mégis, a minap szóba került egy beszélgetés közben a klónozás témája és ismét szembesültem azzal a jelenséggel, amivel korábban találkoztam olyan témák kapcsán, amik eredetileg a molekuláris biológia és biotechnológia belügyeinek számítottak, majd átkerültek a közbeszédbe: sokszor totális a homály még a tájékozottabbak körében is.
Klónozás. A téma annyira snassznak-kommersznek tűnik, hogy már-már cikivé válik írni róla ismeretterjesztő anyagot, olyan szempontból pedig nem vagyok egyszerű helyzetben, hogy ez egyáltalán nem területem. Mégis, a minap szóba került egy beszélgetés közben a klónozás témája és ismét szembesültem azzal a jelenséggel, amivel korábban találkoztam olyan témák kapcsán, amik eredetileg a molekuláris biológia és biotechnológia belügyeinek számítottak, majd átkerültek a közbeszédbe: sokszor totális a homály még a tájékozottabbak körében is.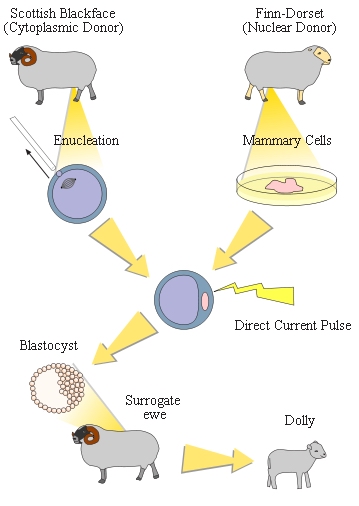
 Ma végülis nem volt erőm megszülni egy megfelelő minőségű cikket az olyan praenatalis diagnosztikai módszerek érdekességeiről, mint az amniocentesis vagy a chorion-boholy mintavétel, viszont kaptok helyette egy saját gyártmányú, háztáji bulvár bullshitet.
Ma végülis nem volt erőm megszülni egy megfelelő minőségű cikket az olyan praenatalis diagnosztikai módszerek érdekességeiről, mint az amniocentesis vagy a chorion-boholy mintavétel, viszont kaptok helyette egy saját gyártmányú, háztáji bulvár bullshitet.  Az elmúlt néhány napban bemutatott hypeok molbiolos fricskázása után kénytelenek lesztek beérni egy kis szájbarágóval. Igen. Algák, gyógyszermellékhatások és az evolúció. Mi köze van ezeknek egymáshoz?
Az elmúlt néhány napban bemutatott hypeok molbiolos fricskázása után kénytelenek lesztek beérni egy kis szájbarágóval. Igen. Algák, gyógyszermellékhatások és az evolúció. Mi köze van ezeknek egymáshoz?  Mint sok más, globális egészségügyi probléma területén, az AIDS gyógyításával kapcsolatban is rendre röppennek fel hírek azzal kapcsolatban, hogy megtalálták a betegség ellenszerét.
Mint sok más, globális egészségügyi probléma területén, az AIDS gyógyításával kapcsolatban is rendre röppennek fel hírek azzal kapcsolatban, hogy megtalálták a betegség ellenszerét. 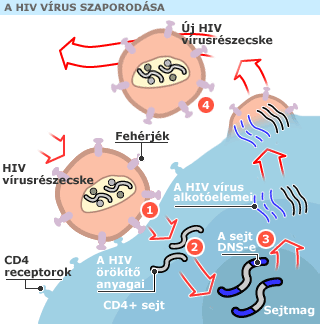
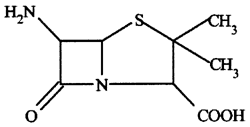 A legkorábban felfedezett, egyes irodalmi források szerint csak a második világháború táján több, mint 150-160 tonna (!!) mennyiségben legyártott pencillin és a most használt penicillin kifejezés alatt valójában egy igen népes vegyületcsoportot értünk, ezen vegyületek közös tulajdonsága a szerkezetükben megtalálható amino-penicillánsav csoport, aminek a tényleges antibakteriális hatás tulajdonítható.
A legkorábban felfedezett, egyes irodalmi források szerint csak a második világháború táján több, mint 150-160 tonna (!!) mennyiségben legyártott pencillin és a most használt penicillin kifejezés alatt valójában egy igen népes vegyületcsoportot értünk, ezen vegyületek közös tulajdonsága a szerkezetükben megtalálható amino-penicillánsav csoport, aminek a tényleges antibakteriális hatás tulajdonítható.  Nem tudom eldönteni, hogy itt valaki most nagyon nagyot akart villantani és ennek érdekében a kutatás valamelyik pontján egyszerűen "meghekkelte a rendszert" (l. alább), esetleg nagyobb szenzációként lett kikommunikálva a tömegsajtóba a hír, ha igaz, differenciálódott, érett bőrsejtekből hoztak létre májsejteket.
Nem tudom eldönteni, hogy itt valaki most nagyon nagyot akart villantani és ennek érdekében a kutatás valamelyik pontján egyszerűen "meghekkelte a rendszert" (l. alább), esetleg nagyobb szenzációként lett kikommunikálva a tömegsajtóba a hír, ha igaz, differenciálódott, érett bőrsejtekből hoztak létre májsejteket.  Növények. A magasabb rendűek közt gyakorlatilag nincs, amelyik képes lenne megváltoztatni a helyét, ezért az evolúcióban kénytelenek voltak menekülés helyett más védekezési stratégiákat kialakítani ellenségeik, a növényevő állatok ellen.
Növények. A magasabb rendűek közt gyakorlatilag nincs, amelyik képes lenne megváltoztatni a helyét, ezért az evolúcióban kénytelenek voltak menekülés helyett más védekezési stratégiákat kialakítani ellenségeik, a növényevő állatok ellen.  Amerika egy perfetisiszta ország, ezt tudjuk jól. Valóságos kultúrája van annak, hogy mindenki perel mindenkit minden szarért. Igazából nem is az a legrémisztőbb, hogy egyesek azért perelnek, hogy megmogyorózzák a konkurenciát, kártérítést csikarjanak ki vagy hasonló financiális érdek vezérli őket. A legijesztőbbek a meggyőződéses hülyék, akik mindenféle szakmaiságot nélkülöző érvekkel hozakodtak elő egy per folyamán, mégis nekik ad igazat a bíróság.
Amerika egy perfetisiszta ország, ezt tudjuk jól. Valóságos kultúrája van annak, hogy mindenki perel mindenkit minden szarért. Igazából nem is az a legrémisztőbb, hogy egyesek azért perelnek, hogy megmogyorózzák a konkurenciát, kártérítést csikarjanak ki vagy hasonló financiális érdek vezérli őket. A legijesztőbbek a meggyőződéses hülyék, akik mindenféle szakmaiságot nélkülöző érvekkel hozakodtak elő egy per folyamán, mégis nekik ad igazat a bíróság.  Nemrég a PLoS Biologyn közölték le, hogy a trippert okozó baktérium fertőzési stratégiája trükkösebb, mint ahogyan azt korábban sejteni lehetett volna.
Nemrég a PLoS Biologyn közölték le, hogy a trippert okozó baktérium fertőzési stratégiája trükkösebb, mint ahogyan azt korábban sejteni lehetett volna.